
抗体应答可维持500天!新型糖基佐剂能否打破mRNA疫苗免疫持久性难题?






本文来自微信公众号:RNAScript,作者:一一
在新冠疫情全球大流行期间,mRNA疫苗充分展现了其模块化平台的极速研发生产能力:只要确定了抗原序列,短短数周就能完成从疫苗设计到规模化量产的全流程。可进入后疫情时代,行业却不得不直面一个现实:仅靠研发速度远远不够。如果未来要持续应对不断变异的病毒,mRNA疫苗始终绕不开三大核心瓶颈:免疫维持时间短、对变异毒株的中和活性不足,以及难以解决的「原始抗原印记」问题。
目前产业界的通用思路就是跟随病毒变异更新抗原,病毒变了,疫苗就跟着换。但这种方式本质上是一场疲于奔命的追赶:当新毒株积累了足够多突变后,人体免疫系统会受「免疫记忆」惯性影响,更倾向于激活最初接触抗原产生的抗体,反而让针对新毒株的免疫效果大打折扣。
前不久,《自然·免疫学》刊登了一项重磅研究,论文题为《一种基于糖基的佐剂可拓展mRNA疫苗的保护广度与持续时间》,该研究提出了一个颠覆传统认知的新方向:mRNA疫苗的下一次升级,不应该只纠结「更换什么抗原」,更应该聚焦「如何精准调控抗原呈递后的局部免疫过程」。
研究团队研发了一种源自真菌的糖基佐剂mannadjuvant(简称MA),将它和mRNA疫苗联合使用。实验结果显示,这款组合方案不仅能明显提升免疫应答的强度和覆盖范围,还能在动物模型中将保护效力延长至惊人的500天,更在临床前模型中首次有效缓解了原始抗原印记对新疫苗效果的限制。这也预示着,mRNA疫苗平台正在从单纯的「抗原递送载体」,迈入深度改造「免疫微环境」的新阶段。
mRNA疫苗为什么还要额外添加佐剂?
在传统疫苗研发中,佐剂的作用很直接:激活先天免疫系统,形成局部炎症环境,以此「推动」适应性免疫系统记住抗原特征。
但放到mRNA疫苗体系中,这个问题就要复杂很多。一方面,mRNA疫苗常用的脂质纳米颗粒(LNP)递送系统本身搭配修饰核苷酸,就自带一定的「自佐剂」效应,可以激活局部炎症反应。所以在行业发展早期,不少观点都认为mRNA疫苗不需要额外添加佐剂。
可随着越来越多临床数据积累,这种内源性免疫刺激的缺陷越来越明显:抗体滴度下降速度快,面对新变异毒株时,生发中心的免疫应答质量达不到要求。能不能通过添加外源佐剂优化mRNA疫苗的免疫质量,已经成为学术界和产业界共同关注的前沿方向。
不过,改造疫苗配方的难度极大。LNP的物理稳定性极差,加入任何外源物质都可能导致颗粒聚集、mRNA泄漏,甚至让递送效率骤降。在不破坏LNP结构的基础上兼容外源佐剂,一直是之前难以突破的工艺门槛。
真菌来源糖基佐剂:从dectin-2到mannadjuvant
本次研究聚焦的佐剂MA,是由白色念珠菌细胞壁提取的甘露聚糖和经典铝盐复配得到的。甘露聚糖是免疫系统识别真菌入侵的核心靶点,能被C型凝集素受体dectin-2精准识别。在此之前,它已经在蛋白质疫苗中被证实有出色的免疫增强效果,但把它应用到mRNA疫苗平台还是首次尝试。
为了解决配方兼容性问题,研究团队做了大量务实的工艺优化。一开始他们尝试先皮内注射佐剂、再肌肉注射mRNA的分步给药方式,但操作太过复杂,不适合推广。为此团队调整了配方,把MA直接和编码新冠病毒刺突蛋白的mRNA-LNP混合,研发出了固定剂量组合的预混肌肉注射方案。
动态光散射检测和mRNA泄漏实验结果显示,混合后24小时内,LNP的粒径没有出现明显变化,mRNA泄漏率不到3%。这说明这款糖基佐剂在物理层面完全适配现有的LNP递送体系,为后续的转化应用打下了工艺基础。
不止提升抗体浓度:抗体应答可维持到500天
在小鼠追踪实验中,WA1mRNA联合MA的组合方案交出了非常亮眼的体液免疫结果。
除了在免疫初期明显提升针对原始毒株和BA.1毒株的中和抗体滴度,最让人惊喜的是它超长的保护持续时间。追踪数据显示,联合接种组小鼠体内抗刺突蛋白IgG水平在接种后500天依然保持在较高水平,而单独接种mRNA疫苗的对照组,抗体水平已经出现了明显衰减。
进一步的机制研究证实,联合接种组小鼠骨髓中,抗原特异性抗体分泌细胞和长寿命浆细胞的数量明显增加。这类定居在骨髓的长寿命细胞,正是人体长期维持稳定抗体水平的核心来源。
这正好击中了目前mRNA疫苗的痛点:在老年人和免疫脆弱人群中,抗体保护效果不到半年就会明显下降。通过调控先天免疫和生发中心反应,把短期的抗体峰值转化为长期的免疫记忆,正是这次技术升级带来的核心突破。
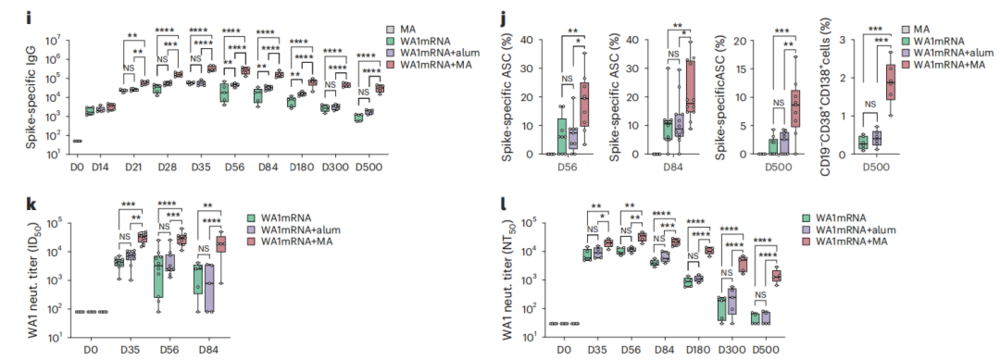
图1.MA可延长mRNA疫苗诱导的体液免疫应答时长
从广谱识别到大动物攻毒验证
除了延长免疫持久性,WA1mRNA联合MA还提升了疫苗对不同变异毒株的识别能力。
在小鼠实验中,研究人员观察到,WA1mRNA+MA可以诱导出更多能够识别BA.4/BA.5和XBB.1.5刺突蛋白的生发中心B细胞和记忆B细胞。血清中和实验也证实,联合接种组对BA.5和XBB.1.5假病毒的中和活性可以维持到接种后第500天,而单独mRNA接种组的中和活性明显更弱。
这个结果说明,MA并不是简单放大已有的抗体反应,而是可能从根本上改变了B细胞反应的覆盖范围和质量。
研究团队还在K18-hACE2小鼠攻毒模型中,验证了这种免疫增强效果能不能转化为实际保护力。接种后第56天,给小鼠通过鼻腔感染BA.5毒株。结果显示,WA1mRNA+MA组小鼠肺部的病毒载量降到了检测不到的水平,肺组织的病理损伤也明显减轻;相比之下,单独接种mRNA疫苗的组仍然能检测到肺部病毒复制,肺组织损伤也更明显。
非人灵长类实验也佐证了这一结论。研究团队在食蟹猴身上对比了单独接种WA1mRNA和联合接种WA1mRNA+MA的免疫效果,发现联合接种组在第180天仍然能诱导出更高水平的针对WA1、BA.5和XBB.1.5的中和抗体。同时,两组食蟹猴的体温、体重和血液指标都没有明显差异,说明在该实验条件下,MA不会明显增加疫苗的不良反应。
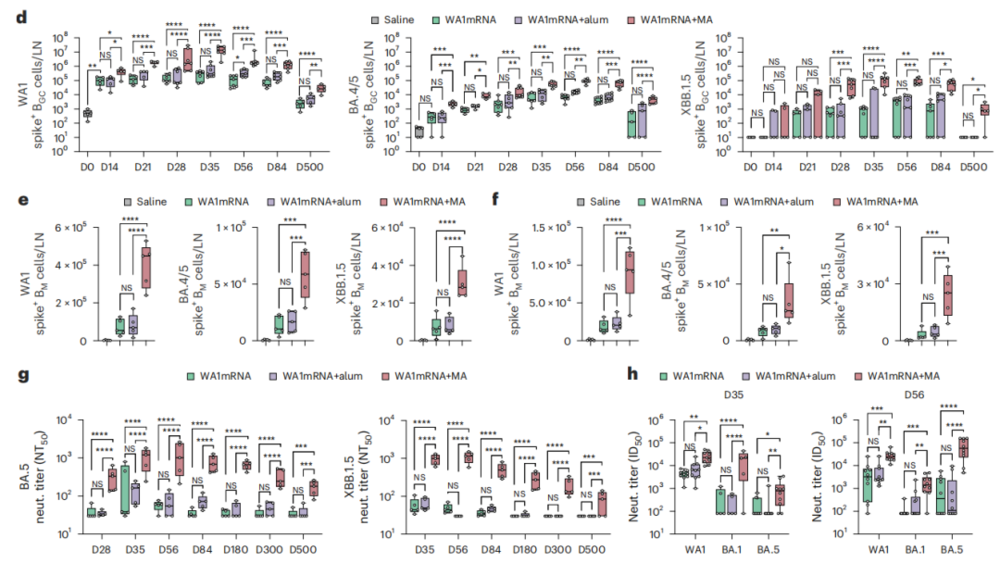
图2.MA拓展了mRNA疫苗诱导的变异毒株识别能力
核心机制:IL-1与I型干扰素重塑局部免疫过程
这项研究最有价值的部分,是它通过时序批量RNA测序,解析了引流淋巴结内形成的时空正反馈免疫过程。
传统的mRNA-LNP疫苗接种后,淋巴结局部的炎症爆发通常在第3天就快速消退,导致B细胞来不及完成充分的分化成熟。但加入MA之后,淋巴结内的细胞因子微环境被彻底改变:
-
早期炎症激活:接种后第1天,MA快速激活了以Nlrp3和Il1a表达为特征的炎性小体反应,提升了IL-1信号水平。
-
中后期干扰素维持:正是因为有早期IL-1信号的铺垫,在接种后第3天到第7天,淋巴结内诱导出了强度高、持续久的I型干扰素和II型干扰素应答。
阻断实验进一步证实,这两条通路并不是独立作用,而是形成了相互促进的正反馈环路——早期的IL-1信号是后续干扰素应答的启动开关,而持续存在的干扰素信号又是维持局部IL-1β水平的关键。
单细胞RNA测序和单细胞BCR测序结果显示,正是这种高质量、长时间维持的细胞因子微环境,明显扩大了生发中心内明区和暗区的B细胞群体,促进了B细胞轻链的体细胞超突变,还降低了单一寡克隆的绝对优势。
这意味着,B细胞在生发中心里经历了更充分的筛选,最终产生了克隆分布更均匀、表位覆盖更广的优质抗体库。VirScan表位分析甚至发现,联合接种诱导出的抗体不仅能识别新冠病毒的全表位,还能结合MERS-CoV和OC43等远缘冠状病毒的部分表位。
有效缓解原始抗原印记问题
对于产业界来说,这篇论文最实用的突破就是破解了原始抗原印记(也叫免疫原性禁锢)的难题。
为了模拟真实人群的免疫背景,研究人员先用原始株mRNA给小鼠做了初次免疫和加强免疫,之后再用变异株XBB.1.5 mRNA做加强接种。结果符合典型的原始抗原印记特征:单独接种XBB.1.5疫苗对新毒株的中和滴度提升非常有限。
但如果在加强针中加入MA后,小鼠针对XBB.1.5的中和抗体水平出现了大幅提升,效果几乎接近直接接种新疫苗的无预存免疫小鼠。
这说明,通过调控生发中心反应和BCR亲和力成熟过程,外源佐剂有可能打破现有免疫记忆的限制,推动免疫系统针对新抗原表位启动高质量的从头免疫应答。
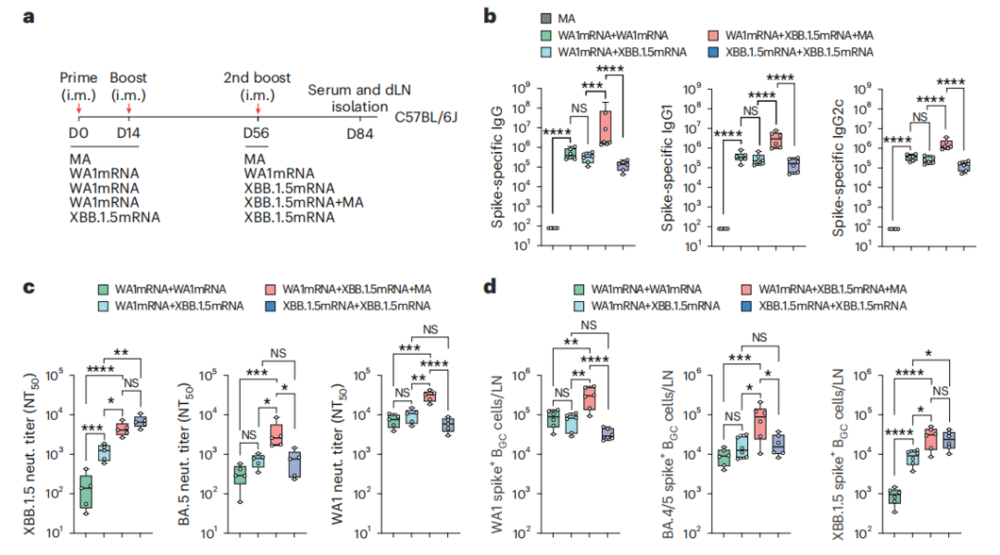
图3.MA缓解了预存免疫记忆对新毒株应答的限制
对产业化落地的冷静思考
虽然这项发表在《Nature Immunology》的研究数据非常扎实,但要真正实现产业化落地,还有几个问题需要解决:
-
补充细胞免疫相关数据:虽然文章主要讨论B细胞和长效抗体,但补充数据显示,联合接种在第500天时,小鼠脾细胞受特异性抗原刺激后分泌IFNγ的水平仍然明显高于对照组。这提示这款佐剂同样可以增强长期TFH辅助和Th1偏向的细胞免疫。但对于完全依赖T细胞的肿瘤新抗原疫苗等应用场景,这种真菌糖基佐剂能不能同样改造肿瘤免疫微环境,还需要针对性的实验验证。
-
平衡反应原性与给药剂量:IL-1与I型干扰素通路的持续激活是一把双刃剑。虽然非人灵长类实验没有观察到明显的全身毒性,但人体对这类强效先天免疫激活剂的耐受范围通常比实验动物更窄。如何在保证「500天长效保护」的同时,把注射局部红肿、疼痛等不良反应控制在三期临床可接受的范围内,是后续剂量优化的核心难点。
-
验证平台普适性:不同企业使用的LNP组分(比如不同的可电离脂质)和mRNA纯化工艺(比如双链RNA残留率)都有差异,这些都会影响内源性免疫背景。MA能不能在不同的LNP体系中保持同样的物理兼容性和免疫重塑效果,还需要更多横向测试验证。
结语:从抗原递送到免疫程序调控
整体来看,这项研究的意义远不止于改良新冠疫苗。它给整个行业展示了一种全新的研发逻辑:
mRNA平台的模块化和研发速度,曾经让我们寄希望于通过「不断更换抗原序列」解决所有问题。但面对高变异、高免疫逃逸的病原体,只靠调整前端抗原序列的优化方式,效果已经越来越差。
MA的价值在于,它成功在现有的mRNA-LNP系统上叠加了一层高效的「先天免疫调控层」。通过激活dectin-2通路,人为构建了一个长效、高质量的淋巴结炎症微环境,精准设计了抗原被免疫系统识别、筛选并形成长期记忆的过程。这种通过外源佐剂重塑免疫程序的思路,或许才是下一代广谱传染病疫苗和治疗性mRNA药物真正走向成熟的必由之路。
本文仅代表作者观点,版权归原创者所有,如需转载请在文中注明来源及作者名字。
免责声明:本文系转载编辑文章,仅作分享之用。如分享内容、图片侵犯到您的版权或非授权发布,请及时与我们联系进行审核处理或删除,您可以发送材料至邮箱:service@tojoy.com